18-20 avril 2025
Pendant 3-4 jours, on a fouillé dans les faluns de Touraine avec la SGAM. Les faluns sont des roches caractéristiques des bassins épicontinentaux peu profonds composées essentiellement de débris coquillers. Les faluns de Touraine se sont déposés dans un bassin qui a été inondé plusieurs fois entre l’Éocène et le Pliocène (les faluns dans lesquels on a prospecté, nous, ont été déposés durant le Miocène moyen). Ils sont surtout connus des amateurs de dents de requins.

Channay
Une fois arrivés, on est tout de suite allé à la carrière-musée de Channay-sur-Lathan. On a trouvé plein de petits fossiles d’invertébrés bien préservés, mais pour les dents de requins, j’étais le seul a en avoir trouvé ; c’était 2 ou 3 petites dents cassées. Puis on se déplace dans une autre carrière, et là on trouve pas mal de dents de requins, mais le soir approche et on a pas beaucoup de temps pour chercher. Je n’ai rien trouvé de très intéressant ce jour ci.


Le deuxième jour, on a exploré une troisième carrière. J’ai trouvé un oursin (différent des scutelles abondantes dans les faluns) et beaucoup de restes de Vertébrés. L’essentiel de mes trouvailles ont été faites sur un remblais qui avait été lavé et déposé par une machine. Il y avait beaucoup de fossiles de Vertébrés à la surface, et je n’était pas le premier à visiter le talus : il y avait des traces de pas d’autres chercheurs. Certains membres du groupe ne trouvent pas de dents ; verdict : on n’est pas dans la bonne carrière. Retour à celle d’avant. La-bas, je tente de trouver un horizon de concentration qui contiendrait beaucoup de dents de requins (on en trouve un peu dans toutes les couches de la carrière, mais leur concentration est faible par rapport à certains talus). Et je trouve un horizon de condensation déposé sur une surface discontinue, teintée d’oxydes de fer et contenant des concrétions de phosphorite : typique des horizons à dents. C’est à ce moment que les fouilles se terminent ce jour là.


Le dernier jour de fouilles, on commence par chercher des calcédoines dans une carrière de calcaire. Une partie du groupe y reste l’après-midi. Moi, je reviens avec quelques autres participants aux dents de requins : j’espère continuer l’exploitation de l’horizon de condensation que j’ai découvert. Mais les fouilles n’ont pas livrées beaucoup de dents, les seules trouvailles étant une demi-dent d’Aetobatus (dent de raie, un assez gros spécimen) et quelques dents plus petites. Bon je me suis pas trop acharné dessus, car on avait pas énormément de temps, et on a fouillé dans un talus qui contenait une quantité phénoménale de dents. Ce talus a visiblement été lavé par la pluie, qui a emporté le sable et laissé les dents. D’autres membres nous rejoignent, tout le monde fait de superbes trouvailles !


Calcédoines



18 mai 2025
La conférence était l’une des nombreuses du festival «Versions Mangiennes» (Mangiennes est une petite ville en France, dans le département de la Meuse). Contrairement à l’année passée, ce n’était pas mon initiative ; l’organisatrice de l’événement m’a invité pour faire une conférence sur la paléontologie.
J’ai préparé un texte et une présentation à projeter avec des photos et quelques explications. J’ai aussi emporté quelques fossiles de ma collection, histoire d’avoir quelques vrais fossiles à montrer. J’ai eu un peu de mal avec le texte : au début, je voulais faire quelque chose de trop compliqué. Je me suis rendu compte que cela rendrait ma présentation trop éloignée et abstraite, et pas grand monde ne risquait d’être intéressé, mais que faire : le plan et une partie du texte étaient déjà prêts. J’ai tenté d’améliorer certaines choses tout en gardant la même structure, mais j’ai vite compris que c’était impossible. Finalement, j’ai complètement changé de structure (en m’inspirant largement de celle de l’année passée) et réécrit tout le texte une semaine avant la date de ma conférence.
J’ai profité de l’occasion de chercher des fossiles dans les environs de Mangiennes. Il se trouve qu’un des participants était aussi un amateur de recherche de fossiles ; on a organisé une petite prospection un matin. Au début, il semblait qu’il n’y avait aucune chance de trouver des fossiles dans cet endroit. Mais, en scrutant un tas de terre sur le bord de route, je vois un minuscule lambeau d’argiles oranges qui n’a l’air de rien d’autre que d’une roche sédimentaire potentiellement fossilifère. Je commence a la gratter avec le marteau de géologue que j’emporte toujours avec moi. L’instinct ne m’a pas trompé : je trouve quelques petits fossiles. Puis, je tombe sur une pièce assez exceptionnelle : un morceau de carapace fossilisée d’un crustacé. On continue nôtre chemin, et on arrive sur des tas du même argiles, mais beaucoup plus grands. On ne sait pas d’où ils viennent, probablement d’une carrière ou d’un chantier local. C’est là que je passerai tout l’après-midi. J’arrive à situer ces argiles dans le Jurassique moyen.



La conférence avait de nouveau comme sujet la paléontologie et les fossiles en général. Elle s’est passée un peu moins bien que j’avais prévu : il me restait peu de temps de préparation à cause des changements dernière minute. J’avais aussi demandé un projecteur, mais tout ce qu’on a trouvé sur place était un grand écran d’ordi. J’ai improvisé une petite exposition avec des fossiles de ma collection, et j’ai naturellement ajouté ceux que je venais de trouver.
Mon travail au MGL consistait à transférer des informations sur des fossiles conservés au musée depuis des fiches manuscrites vers un catalogue numérique. J’ai entré environs 400 spécimens, principalement des fossiles du canton de Vaud.
Début juin, j’ai pu visiter les fonds du MGL à l’UNIL avec Antoine Pictet (conservateur du musée).
Pendant le camps d’été de cette année, on a visité une mine avec fluorite + barytine et les haldes de Wittichen et Grube Clara. Pour moi, le lieu le plus intéressant était Wittichen, les talus de mines abandonnées qui contiennent, entre autre, des minéraux uranifères. Grube Clara est très touristique, mais on y trouve vraiment une diversité de microminéraux impressionnante. J’espère pouvoir faire des photos de mes trouvailles, car bientôt j’aurais du nouveau matériel (les cristaux sont touts petits et nécessitent des objectifs à fort grossissement). Les minéraux uranifères devraient donner de beaux résultats en ultra macro avec fluorescence UV.

Microminéral avec une croûte d’argent natif. Petite Halde (Grube Clara). Largeur de l’image : env. 3mm.
Cette année, je me suis beaucoup intéressé aux microfossiles, qu’on peut trouver dans un grand nombre de roches mais seulement avec un microscope, car ils mesurent entre 0,1 et 2 mm (d’ailleurs j’ai, depuis le début de l’année, un microscope puissant qui a rendu la recherche plus efficace). Au printemps, j’ai proposé à la SGAM d’organiser une exposition de microfossiles au local. Le local de la SGAM m’a semblé un bon endroit pour cet événement, car il y avait 4 loupes binoculaires ; parfait pour observer les microfossiles. Seulement, à ce moment là, l’observation avec ces outils était peu pratique, non pas à cause de leur qualité qui était au top pour les 4 modèles, mais parce qu’il fallait faire quelques réparations et un entretien des mécanismes, sans parler du nettoyage, sans lequel l’image était légèrement trouble. J’ai déjà eu le temps de restaurer deux loupes binoculaires. J’espère pouvoir annoncer la date de l’exposition dans quelques mois, et je vais continuer à la préparer en 2026.
Cette publication démarre une série de 5 articles sur un groupe particulier de fossiles : les ammonites. Ce sont des coquilles de céphalopodes appartenant à un groupe éteint: la sous-classe Ammonoidea. Ce ne sont donc évidemment pas des «escargots de mer», même si la forme des coquilles ressemble un peu à celle des gastéropodes : comme tous les céphalopodes, les ammonites possédaient une tête avec 2 yeux bien développés et plusieurs tentacules ; elles ne rampaient pas sur le fond marin mais se déplaçaient en nageant (necton). Les premières ammonites sont apparues il y a près de 400 millions d’années au Dévonien. Ce groupe était dominant dans les mers et les océans pendant près de 334 millions d’années avant de disparaître complètement il y a 66 millions d’années. Les ammonites ont une grande importance en paléontologie : leurs fossiles sont parmi les plus abondants, surtout dans les roches du Mésozoïque. Les coquilles spiraliformes sont répandues dans un grand nombre de roches, comme par exemple les calcaires et les marnes du Jura (Jurassique-Crétacé), où on trouve aussi de nombreuses plaques appelées Aptychi, qui correspondent à la mâchoire inférieure des ammonites.

Ammonites de l’Albien (Crétacé inférieur) du Jura, sur le site de découverte.

Ammonite Quenstedtoceras flexicostatum. Oxfordien inférieur, Jura, Vaud. Voir dans la gallerie

Laevaptychus sp., élément de la mâchoire inférieure d’une ammonite (Aspidoceratidae indet.). Oxfordien inférieur, Jura, Vaud. Voir dans la gallerie
Le terme «ammonite» était déjà utilisé dans l’Antiquité pour désigner ces fossiles. Il est lié au dieu égyptien Amon. Celui-ci était souvent représenté avec des cornes de bélier, qui ressemblent aux coquilles d’ammonites.
 Ammonite Cantabrigites aff. cantabrigense. Albien (Crétacé inférieur), Jura. Voir dans la gallerie
Ammonite Cantabrigites aff. cantabrigense. Albien (Crétacé inférieur), Jura. Voir dans la gallerie
Pendant longtemps, il n’y avait pas d’explication scientifique pour l’apparition des fossiles. On ne les liait que rarement à des organismes vivants, sinon on pensait qu’ils étaient capable d’apparaître spontanément dans la roche. Le premier à identifier les ammonites comme des céphalopodes est Robert Hooke (1635-1703). Hooke a comparé des fossiles d’ammonites qu’il a trouvé avec Nautilus, un céphalopode actuel avec une coquille en spirale.
Les Nautilidae modernes utilisent leur coquille pour assurer une flottabilité neutre à différentes profondeurs. Les tours intérieurs de leur coquille sont séparés en sections internes, appelées chambres hydrostatiques, par des cloisons. La coquille se termine par la loge d’habitation, une grande portion de la coquille non cloisonnée qui contient le corps du mollusque et se termine par une ouverture. La partie cloisonnée, elle, est appelée phragmocône. Les nombreuses sections du phragmocône contiennent de l’eau et du gaz. Toutes les sections sont transpercées par un tube appelé siphon, dont la fonction est de modifier la quantité d’eau et de gaz dans chaque chambre hydrostatique. Grâce à ce mécanisme, les nautiles peuvent remonter vers la surface en diminuant la quantité d’eau et en augmentant la quantité de gaz dans le phragmocône, et vice versa.
 Nautilus pompilius actuel. Source de l’image.
Nautilus pompilius actuel. Source de l’image.
 Coupe d’une coquille d’un Nautilus actuel. Source de l’image.
Coupe d’une coquille d’un Nautilus actuel. Source de l’image.
En observant leurs coquilles fossiles, R. Hooke a remarqué que les ammonites avaient une organisation similaire. Les cloisons, bien que beaucoup plus complexes que chez les nautiles, divisent la coquille en chambres hydrostatiques, et là où les derniers tours de la coquille sont préservés, une portion de la coquille est libre de cloisons. Le siphon est préservé chez certains spécimens d’ammonites. Robert Hooke ne connaissait pas la fonction du siphon, il l’a néanmoins remarqué sur quelques uns de ses spécimens fossiles.
 Ammonite Dorsoplanites dorsoplanus. Moscou, Jurassique supérieur, env. 149 millions d’années. Remarquez le siphon sur la photo de droite. Voir dans la galerie
Ammonite Dorsoplanites dorsoplanus. Moscou, Jurassique supérieur, env. 149 millions d’années. Remarquez le siphon sur la photo de droite. Voir dans la galerie
Avec ses observations sur les ammonites et d’autres fossiles, Robert Hooke énonce plusieurs hypothèses. Il développe l’idée de fossilisation par remplacement des restes organiques par des substances minérales présentes dans l’environnement. Il observe aussi qu’un grand nombre de fossiles marins sont présents très loin des mers, d’où il conclut que des mers, qui existaient autrefois, ont disparu à cause de changements dans le relief de la surface terrestre. Enfin, il démontre que certains fossiles appartiennent à des groupes d’animaux éteints, et il suppose qu’une évolution des formes vivantes peut avoir lieu.

Ammonites figurées par Robert Hooke (1).
Les ammonites sont des fossiles utiles en géologie. Leur classification bien développée et leur évolution rapide permet d’utiliser différentes espèces d’ammonites comme repères temporels pour dater des couches de roche sédimentaire. Les ammonites donnent aussi des indications sur là paléogéographie marine, telle que les voies marines ou les courants marins, grâce à la migration de différentes espèces.
 Source: Mironenko, A.A., 2015: Wrinkle layer and supracephalic attachment area: implications for ammonoid paleobiology. Bulletin of Geosciences 90(2): 389–416
Source: Mironenko, A.A., 2015: Wrinkle layer and supracephalic attachment area: implications for ammonoid paleobiology. Bulletin of Geosciences 90(2): 389–416
L’image ci-dessus est une reconstruction d’une ammonite. Cette reconstruction représente une vision moderne de ce que sont les ammonites selon les connaissances actuelles en paléontologie. J’aurai encore l’occasion de discuter de l’exactitude de différentes reconstruction dans un autre article.
L’organe allongé sous la tête du mollusque est appelé hyponome, parfois aussi «funnel» (en anglais) ou siphon (ne pas confondre avec le siphon du phragmocône dont il est question plus haut). C’est un organe de locomotion commun à tous les céphalopodes actuels. La locomotion implique la cavité du manteau (mantle cavity sur le dessin ci-dessous), qui se remplit d’eau. L’eau est ensuite éjectée de la cavité par l’hyponome avec force, ce qui crée une force de propulsion qui pousse le mollusque en arrière.
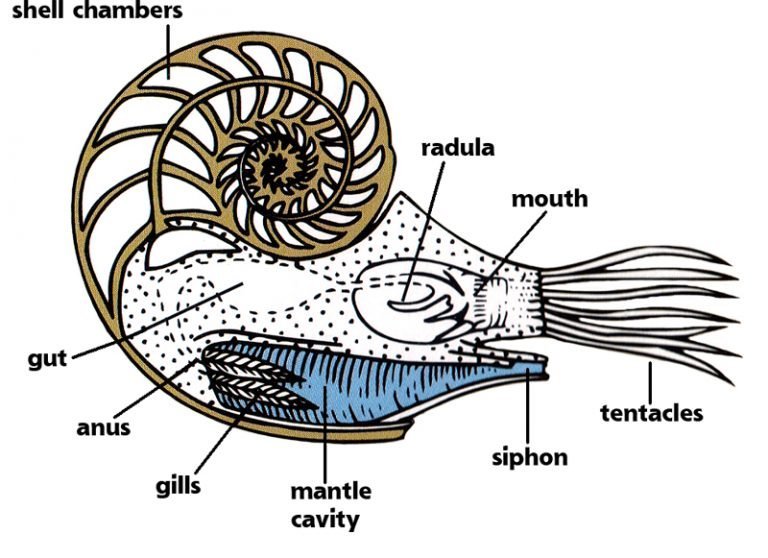
Source de l’image
On pense que le mode de locomotion par propulsion était le principal chez la majorité des ammonites, et leur permettait de se déplacer assez rapidement. Il est toutefois possible que certaines ammonites utilisaient d’autres moyens de locomotion ; quelques espèces possédaient peut-être des membranes entre leur tentacules qui leur permettaient de nager en ramant (ce sont des spéculation basées sur les calmars et les pieuvres actuels, on n’a retrouvé aucun fossile de tentacules qui puisse les vérifier). Enfin, certaines ammonites n’étaient pas adaptées à la locomotion active et restaient suspendues dans l’eau, en se laissant transporter par les courants marins.
Evolutivement, les ammonites sont plus proches de Coleoidea que de Nautiloidea. Ce qui peut paraître bizarre, surtout que j’ai dit juste avant que c’est la comparaison avec les nautiles qui a permis de comprendre que les ammonites étaient des céphalopodes. En réalité, c’est l’évolution divergente des coleoides qui prête à confusion : les coleoides ont acquis d’abord une coquille interne, voir par exemple les bélemnites. Puis, cette coquille a perdu son importance, ce qui a d’abord entraîné la perte du phragmocône chez plusieurs groupes tels que les calmars et les seiches, puis enfin la disparition de la coquille chez les poulpes. Les nautiles, eux, ont gardé la coquille externe avec phragmocône. Une partie de la ressemblance avec les coquilles des ammonites est due à des caractères primitifs communs, une autre à la convergence. L’origine des ammonites ainsi que leur proximité avec les coleoides actuels méritent d’être discutée plus en détail, ce qui sera fait dans le prochain article.

Fossile de bélémnite (Cephalopoda; Coleoidea), Jurassique. La coquille, consistant du seul phragmocône (partie grise) est situé à l’intérieur du corps du mollusque.
Ne manquez pas : Paléobiologie et évolution des ammonites. Partie 2
- Hooke R., Discourse on Earthquakes. The posthumous works of Robert Hooke p. 279-289
- Ogura A., Yoshida M., Moritaki T., Okuda Y., Sese J., Shimizu K.K., Sousounis S., Tsonis P. A. Loss of the six3/6 controlling pathways might have resulted in pinhole-eye evolution in Nautilus. // Scientific Reports. 2013. Vol. 3. art. 1432.
L’année 2024 est presque écoulée, et je marque cette fin d’année avec un petit compte-rendu de mes activités paléontologiques. Celles-ci comptent des fouilles très productives, la découverte de nouveaux sites fossilifères et bien d’autres choses.
Un des événements important est bien sur la création de ce site, ainsi que de la galerie de collection gallery.lsglab.org, non sans l’aide de Julien Mabillard. Je dois remercier Julien pour avoir mis en place une solution très pratique pour mon utilisation. Première publication sur la galerie : 19 janvier 2024, sur lsglab.org : 26 avril 2024. Je compte continuer le développement du site dans l’année à venir, et j’ai déjà des plans pour quelques grands articles sur la paléontologie et la théorie de l’évolution.
Très mouillé, une cascade descend la paroi de roche, ce qui n’empêche pas de trouver de bons spécimens.

Canton de Vaud, Jurassique supérieur (étage Oxfordien).

Brachiopodes, ammonites Perisphinctidae, éponges, fragment de lys de mer.
De nouveau mon site dans le canton de Vaud. Cette fois, j’ai étudié d’autres niveaux qui n’étaient pas visibles à la surface. Il a fallu creuser pour les exposer.

Selon quelques information vagues, on y avait trouvé des dents de requins. Comme c’était de l’argile, j’ai pris mon tamis et de l’argile dans un sac que j’ai transporté jusqu’à une rivière qui coule à côté de l’endroit de fouilles. Le résidu tamisé contenait beaucoup de débris, dont des cailloux et des morceaux de bélemnites, ainsi que des petites concrétions de phosphorite. Il aurait été dur de repérer des dents dedans, et j’ai commencé à douter qu’il y en avait… J’ai pensé bien des fois à vider le reste du sac et abandonner, mais, pour être sûr du résultat, j’ai continué à tamiser jusqu’au bout. Et, vers la fin, une forme allongée familière est apparue dans le tamis. Je saisis d’abord un bélemnite qui se trouvait à côté, mais je comprend vite que ce n’est pas ce que je cherche. Je continue à fouiller dans les cailloux pour retrouver l’objet perdu, et je retrouve cette…

… petite dent de plésiosaure

Puis une dent de requin a rejoint la collection, confirmant la richesse de ce site en vertébrés.
Ce site est devenu le numéro 1 cette année. J’y suis revenu de nombreuses fois dans l’espoir de trouver d’autres dents de reptiles marins, mais malheureusement j’ai été déçu de ce point de vue là. C’est néanmoins un site très intéressant, autant pour les dents de requins que pour certains invertébrés qui ont été trouvés plus tard.
J’y étais avec un paléontologue du musée et un archéologue. On a promis à l’archéologue de lui donner les clous rouillés si on en trouve pendant les fouilles. Les fossiles trouvés sur place sont au musée. J’ai pris un échantillon de sédiments qui a donné quelques dents de poissons et coquilles.
Retour au Jurassique vaudois, avec un ami. On a commencé par tamiser, mais il n’a pas trop apprécié, je cite : «Max, je me les gèle, et en plus on ne trouve rien». Le tamisage ici, ce n’est pas comme à Shatsk, il demande de la patience. Je lui ai montré l’endroit un peu plus haut où j’avais fouillé la première fois, où on trouve des ammonites et des éponges. Il a trouvé quelques spécimens et surtout fait la sieste au soleil. J’ai continué à tamiser seul, puis on est rentré. Résultat d’une journée de tamisage intensif : quelques Aptychi, un Rhyncholite et une seule dent, mais une Hexanchiforme (qui sont extrêmement rares).
Week-end de prospection dans le Vercors (Crétacé supérieur, étage Campanien). Le premier jour : rien ! On est revenu un peu sur nos pas et placé la tente à un endroit assez sympa vers un ruisseau. Le deuxième jour, j’ai recommencé à prospecter, et il s’est avéré qu’une paroi exposait un niveau à microfossiles juste en face du campement, de l’autre côté de la route ! Le tri des échantillons de roche continuera l’année prochaine.

Sortie dans une carrière avec la SGAM, Oxfordien (Jurassique supérieur). A part les cristaux de pyrite et de célestine, on y trouve des fossiles, et la composition en fossiles de chaque étage de la carrière est différente.

Recherches dans le Miocène de l’Ain
Ma première exposition paléontologique, qui a pris 3 mois à préparer, a enfin eu lieu. J’ai fait une présentation d’environs 20 minutes. Le compte-rendu complet à déjà été publié ici. Il y a eu une distribution de dents de requins du Miocène (20 millions d’années) à la fin.
Je vais probablement répéter ce genre d’événements. Une conférence paléontologique est prévue le printemps prochain dans le département de la Meuse.






Une prospection dans le canton de Vaud, pas très réussie. D’abord découverte tout à fait inattendue d’argiles aptiens oranges, niveaux assez rares, comme ceux de l’échantillon du 09.03. Mais je n’ai pas réussi à trouver les horizons fossilifères, et mon échantillon de cette fois-ci était vide. Puis, quelques recherches dans un bord de route de l’oxfordien, jusqu’à ce qu’un paysan vienne me dire que mon trou de 15x15x15 cm risque de provoquer une extinction de masse de l’herbe sur son champ, situé au moins 5 m plus haut. Donc, j’ai pris un échantillon d’argile et je suis parti… Résultats : j’ai trimballé 2 gros sacs d’argiles, côté fossiles : 3 petites ammonites, assez bien conservées, mais rien d’exceptionnel, et un Laevaptychus fragmenté. Aucune dent, qui étaient un peu le but de cette prospection.

17.08 – Mine de phosphates abandonnée (France)
Cette mine est riche en fossiles de l’albien, principalement des moules internes de mollusques, mais aussi des dents de requins. On pouvait y trouver des bons spécimens à une époque, à ce qu’on m’a dit. Mais la longue collecte excessive et surtout l’ignorance des fouilleurs envers des techniques efficaces a appauvri le site. J’y ai trouvé très peu de dents complètes (les racines sont de toute façon mal préservées dans cette formation). J’y suis retourné encore une fois plus tard dans l’année, avec un résultat encore moins bon pour les dents, mais j’ai par contre trouvé des fragments de crustacés : des pinces, mais aussi un élément de carapace.


La saison étant terminée, je souhaite une bonne année 2025 à tout le monde !
Certaines couches de roche contiennent un grand nombre de fossiles de vertébrés de taille microscopique : moins de 2 mm. Ce sont des petites dents, écailles, fragments de mâchoires… Ils sont souvent très différents des macrofossiles trouvés dans la même couche, et représentent donc des informations d’une grande valeur paléontologique. Pour pouvoir récupérer les microfossiles, il faut commencer par désagréger la roche. Avec une roche calcaire, ça se fait par bains d’acide. Je vous présente la méthode simple et efficace que j’utilise pour extraire des microvertébrés d’une roche calcaire.
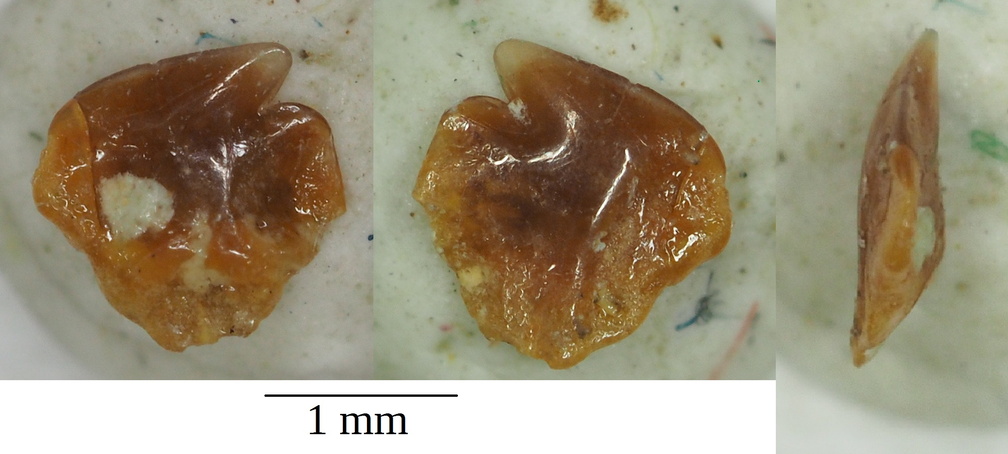
Cretascymnus sp. micro dent de requin
Les morceaux de roche calcaire collectés sur le terrain sont placés dans un bain de vinaigre de table (acide acétique 5-10%) dilué dans de l’eau contenant de l’acétate de calcium en proportion de volume eau:vinaigre:acetate de calcium 8:1:1. L’acétate de calcium peut être obtenu par la dissolution de calcaire dans de l’acide acétique (on voit d’ailleurs souvent des cristaux blancs d’acétate de calcium sur des morceaux de roche dans de l’acide acétique usé). Laisser la roche dans le bain pendant quelques heures, puis simplement rajouter un peu de vinaigre dans la solution usée. Une fois la roche désagrégée, on obtient un résidu de minéraux insoluble (quartz, glauconie etc.) et microfossiles, prêt à être criblé et trié. Le bain usé peut être récupéré et séché pour obtenir l’acétate de calcium.

Cristaux d’acétate de calcium

Cristaux d’acétate de calcium : macrophoto
Cette méthode est très avantageuse pour la préservation de microfossiles, car la présence d’acétate de calcium permet de
- Imprégner la roche avec l’acide. L’acide réagit non seulement à la surface, mais aussi dans l’épaisseur de la roche, et dissout le ciment, plus sensible, sans attaquer les fossiles.
- Eviter toute corrosion des restes de microvertébrés grâce à la présence d’ions de calcium Ca++, qui vont empêcher la réaction du phosphate de calcium (qui compose les microvertébrés) avec l’acide.
Cette méthode m’a même permis d’extraire des foramenifères calcaires d’un grès à ciment calcaire, quoique un bon nombre a été corrodé par l’acide.
Si vous voulez récupérer des fossiles calcaires avec cette méthode, je recommande de récupérer le résidu chaque fois avant de remettre de l’acide, pour diminuer l’exposition des fossiles à l’acide.
Le 25 mai 2024 a eu lieu ma première exposition de fossiles. Pendant 2 ans, mes spécimens étaient conservés chez moi dans des boites (et jusqu’en 2022, à Moscou, j’avais une chambre-musée où j’exposais certains fossiles). En mars, mon père ma dit qu’on pouvait me prêter une salle, et donc que je pouvais organiser une exposition.

Les spécimens de ma collection à Moscou, 2019.
J’ai commencé à préparer mon exposition déjà début mars. J’ai tout de suite préparé les fossiles que j’allais exposer et écrit le texte de la présentation. J’ai apporté quelques corrections à ce texte en mai. Mais le temps de préparation manquait à cause de différentes choses qui n’avaient rien à voir avec l’exposition, mais qui devaient être faites et qui prenaient la majorité de mon temps. La date de l’exposition a finalement été fixée au 25 mai. Je pense que j’aurais dû commencer la préparation encore plus tôt pour me concentrer sur l’organisation les dernières semaines, ce qui aurait permis d’éviter certains problèmes survenus au dernier moment. J’ai par exemple indiqué l’adresse du bâtiment d’à côté sur l’affiche de l’exposition – 9 au lieu de 11. Si j’avais eu plus de temps, j’aurai relu 10 fois l’affiche avant de l’envoyer.
J’ai annoncé l’exposition à mes connaissances, et aussi demandé à un prof d’envoyer l’affiche de l’exposition à toute l’école par mail, ce qu’il a fait.
J’ai exposé les fossiles que j’ai trouvés et conservés dans ma collection : des dents de requins (du Carbonifère, Jurassique, Crétacé et Néogène), des os de reptiles du Mesozoïque, des ammonites du Jurassique et du Crétacé, des bivalves, des gastéropodes, des brachiopodes, des bryozoaires et certains autres fossiles. Je les ai disposés dans des vitrines prêtées par la SGAM. Certains fossiles avaient des étiquettes avec des informations détaillées. Tout le monde connaît les requins, mais peu de gens ont vu des brachiopodes ou des bryozoaires.

J’ai disposé les spécimens dans deux vitrines.
En réalité, il aurait fallu avoir plus de vitrines pour mieux voir les spécimens et les étiquettes.






Os de nageoire (os du poignet et phalanges) d’Ichthyosaure

Bryozoaires (Carbonifère) et leur étiquette détaillée





Au milieu de l’exposition, il y a eu une conférence sur la paléontologie. Elle était divisée en 5 parties:
- Introduction. Qu’est-ce que la paléontologie. Comment se forment les fossiles.
- L’histoire géologique de la Suisse. J’ai parlé des mers et océans anciens de la Suisse: l’océan Téthys et le bassin d’avant-pays Nord-Alpin, car ce sont précisement les roches d’origine marine qui contiennent le plus de fossiles.
- Partie sur les expéditions, où et comment trouver des fossiles.
- L’intérêt scientifique des fossiles. Partie sur les empreintes de muscles sur les fossiles d’ammonites, l’évolution des requins, la biostratigraphie, la détermination des conditions de l’environnement grâce aux fossiles.
- Les adresses des sociétés géologiques et paléontologiques de Suisse.
J’aurais dû aussi faire une partie sur le dégagement des fossiles, mais ce sera pour une autre fois.


Il y avait près de 15 personnes qui ont assisté à la présentation. Tout le monde avait l’air très intéressé. En tout, il devait y avoir 20-25 personnes à l’exposition, dont 3 personnes de mon école. L’un d’eux était très impressionné par des dents de requins de 330 millions d’années. Je dois dire que l’exposition s’est bien passée, et c’est toujours bon pour les fossiles de sortir des boites.s
Leonid Sergeyevich Glickman était un paléontologue spécialiste de l’évolution des requins. Ses recherches ont permis de révolutionner la classification des Elasmobranches. Il a développé de nouvelles méthodes de recherche basées sur l’étude de l’appareil dentaire, qui ont conduit à de remarquables avancements dans l’étude de l’évolution des requins.

Leonid Glickman est né le 23 janvier 1929 à Leningrad. Son père – Serguei Abramovitch Glickman – était un chimiste reconnu. En 1939, Glickman et ses parents déménagent à Kiev. Sa mère meurt un peu avant la guerre. En 1941, Leonid Glickman et son père sont évacués à Tachkent. A la fin de la guerre, ils s’installent à Saratov, où son père est nommé chef de la faculté de chimie colloidale de l’Université de Saratov. A l’âge de 18 ans, L. S. Glickman termine l’école et poursuit ses études à la faculté biologie et sols de l’Université de Saratov.
La région de Saratov est connue pour ses dépôts du Crétacé supérieur qui abondent en dents de requins fossiles. Déjà à la fin de l’école, Leonid Glickman s’intéresse aux Elasmobranches et commence à collecter des dents de requins. Pendant ses études à l’Université de Saratov, il rassemble une collection de quelques dizaines de milliers de fossiles de vertébrés dans les dépôts du Crétacé de la région de Saratov : requins (dents), chimères (dents et os), poissons osseux (dents, os, écailles), ichtyosaures (vertèbres), plésiosaures (dents et os), et un fragment de mâchoire et un squelette partiel de ptérosaures.
L. S. Glickman prend contact avec L. I. Hozatskiy, paléontologue spécialiste des tortues à l’Université de Leningrad. L. I. Hozatskiy l’aide à passer à la faculté biologie et sols de l’Université de Leningrad. En 1950, Glickman déménage à Leningrad et y poursuit ses études. Il les termine en 1952, avec un travail de diplôme intitulé «Les vertébrés marins du Crétacé supérieur des rives de la Volga vers Saratov», qui servira de fondement pour son premier article scientifique [1].
L. S. Glickman cherche à continuer ses recherches dans le domaine de la paléontologie des Elasmobranches. Mais, au début des années 50, c’est le pic de l’antisémitisme en URSS et, malgré ses études brilliantes, il a du mal à trouver du travail avec un nom de famille juif. Il n’est engagé en 1952 qu’en tant que secrétaire au Musée de géologie A. P. Karpinskiy. Il réussit à prendre part à deux expéditions en 1953 (expédition du Laboratoire des Aeromethodes à partir du 2 mai et expédition Caspienne 7 mai – 2 octobre). Puis, en 1954, sur la demande de l’Institut de géologie de l’Université de Saratov, Glickman réalise un travail de recherche sur les requins du Crétacé supérieur (Glickman, 1954). Pour le réaliser, il récolte des dents de requins dans des expéditions dans la région de Saratov et les régions voisines. En 1954, Glickman est engagé en tant que chercheur au Musée A. P. Karpinskiy. Il publie de nombreuses recherches sur l’évolution des requins. En même temps, il commence à préparer sa thèse de candidat*. Le 25 décembre 1958, il défend sa thèse «Sur la classification des requins» avec succès et reçoit le titre de candidat en biologie. Cette thèse deviendra le fondement de la monographie «Les requins du Paléogène et leur valeur stratigraphique» [5], une révolution dans la classification et l’étude des requins. En 1964, il publie également la rubrique «Elasmobranchii» dans les «Fondements de la paléontologie», un ouvrage de référence pour les paléontologues en URSS [6].

L. S. Glickman avec le crâne d’un Carcharhinus actuel, années 60.
Pendant cette période L. S. Glickman récolte de nombreux spécimens de dents requins fossiles. Glickman étudie la stratigraphie du Crétacé et du Cénozoïque de la région de Moscou, de la Volga, du Khazakhstan, de l’Asie centrale, et rassemble une collection détaillée de dents de requins d’horizons differents. La plupart de ces expéditions ont été payées par Glickman lui-même.

L. S. Glickman en expédition au Kazakhstan, années 60.
En 1970, Leonid Glickman déménage à Vladivostok, où il travaillera pendant 10 ans à l’Institut de biologie marine (IBM). Il s’attend à pouvoir étudier les requins actuels, mais n’en a pas vraiment la possibilité. Il se consacre à l’étude des salmonidés, et travaille également comme directeur de recherches et consultant auprès d’autres chercheurs en biologie des vertébrés, ou bien en biostratigraphie. Glickman prépare aussi sa thèse de docteur «Tendances dans l’évolution des requins lamniformes». Il tente d’obtenir le titre de docteur en géologie en 1972 à la faculté de géologie de l’Université de Moscou. N’y étant pas parvenu, il refait une tentative en 1974 à l’Institut de paléontologie de Moscou. Cette fois, il cherche à obtenir le titre de docteur en biologie. Il n’y parvient pas non plus, malgrés une pré-défence réussie. Un des jurés, V. V. Menner, recommande d’améliorer la thèse et en 1976 Glickman cherche à obtenir le même titre à l’IBM, avec le support de V. V. Menner, mais, au même moment, Glickman part en expédition au Kamtchatka pour une demi-année. Il n’entreprendra plus de tentatives de défendre sa thèse. En 1980, elle est publiée sous le titre «Evolution des requins lamniformes du Crétacé et du Cénozoïque» [7].
A la fin des années 70, L. S. Glickman ne s’entend plus avec ses chefs à l’IBM. Il quitte Vladivostok pour revenir à Leningrad. En même temps, sa collection est transférée au musée Darwin à Moscou, où elle se trouve aujourd’hui. Etant très pauvre, Glickman doit travailler comme garde dans une usine, et a une fracture de la hanche qui n’a pas été soignée. Malgré cela il publie toujours des travaux (Glickman et Zhelezko, 1985, Glickman et Dolganov, 1988a, 1988b, Glickman et Averianov, 1998 et autres).
En 1999, Glickman participe à sa dernière expédition paléontologique au Kazakhstan. Il meurt peu après, le 31 Janvier 2000, à l’âge de 71 ans.
* Candidat - titre attribué en URSS et encore actuellement en Russie aux chercheurs sous remise d’une thèse. Le titre de candidat précéde celui de docteur
Dès le début de ses études, Glickman choisit la paléontologie des Elasmobranches comme spécialité. Il se concentre sur l’étude de leur évolution, et cherche à développer de nouvelles méthodes de recherche. Il rassemble une grande quantité de dents de requins et étudie des restes de requins actuels conservés dans des musées. L’étude de ce matériel lui permet de tirer des conclusions importantes sur la systématique et l’évolution des requins.
Glickman a rassemblé une collection de dents de requins de près de 200 000 exemplaires [4], qui compte des exemplaires fossiles d’Europe de l’est et d’Asie centrale récoltés par Glickman lui-même ou par d’autres scientifiques, ainsi que des dents de requins remontées du fonds des océans Indien et Pacifique par les stations de recherche «Vitiaz», «Ob» et «Lomonosov». Il y incorpore aussi les collections de A. S. Rogovitch, I. F. Sinzov et d’autres scientifiques, qui ont été sauvées à la liquidation du musée de géologie A. P. Karpinskiy. La collection est conservée au musée Darwin à Moscou.

L. S. Glickman avec sa collection de dents de requins fossiles, années 50.
En 1970-1980, Glickman a étudié les salmonidés et arrive à faire quelques hypothèses sur leur évolution.
Leonid Glickman a publié 4 monographies (Glickman, 1964a, 1964b, 1980, Glickman et al., 1987) et 33 articles scientifiques, ainsi que des œuvres de vulgarisation. Le concept principal de Glickman est sa classification des elasmobranches. Il a remarqué des différences fondamentales entre l’organisation du neurocrâne [7], des mâchoires et de l’appareil dentaire [5] des requins lamniformes et celui des autres elasmobranches. De plus, les dents des requins lamniformes sont composées d’ostéodentine (tissu ressemblant à de l’os), alors que les dents des autres requins ont une couche d’orthodentine sous l’émail, et un canal pulpaire (Fig. 1). Ceci conduisit Glickman à éloigner les lamniformes des autres requins et raies actuels, et les rapprocher des Elasmobranches éteints ayant des dents composées d’ostéodentine. Il divise ainsi les Elasmobranches en deux infraclasses : les Ortodontes et les Osteodontes, division basée sur l’histologie dentaire [5]. Cette division se confirme par la similitude d’anatomie craniale que presentent les Carcharhini (voir partie “Classification des Elasmobranches d’après Glickman, 1964a”) avec les Xenacanthes (requins d’eau douce du paléozoïque que Glickman supposait être l’ancêtre direct des Carcharhini), et l’organisation similaire du neurocrâne de Chlamydoselachus[7].
Sous-classe Еlasmobranchii
- Infraclasse Orthodonta
- Supérordre Cladoselachii
- Ordre Cladoselachida
- Fam. Сladoselachidae
- Fam. Denaeidae
- Ordre Сladodontida
- Fam. Сladodontidae
- Fam. Symmoriidae
- Fam. Tamiobatidae
- Supérordre Xenacanthi
- Supérordre Polyacrodonti
- Supérordre Chlamydoselachii
- Supérordre Carcharini
- Ordre Hexanchida
- Subordre Hexanchoidei
- Subordre Heterodontoidei
- Ordre Squatinida
- Subordre Echinorhinoidei
- Subordre Squaloidei
- Fam. Squalidae
- Fam. Dalatiidae
- Fam. Cetorhinidae
- Subordre Squatinoidei
- Subordre Ginglymostomatoidei
- Subordre Pristiophoroidei
- Subordre Rajoidei
- Supérfam. Rhinobatoidea
- Fam. Rhinobatidae
- Fam. Asterodermatidae
- Fam. Platyrhinidae
- Supérfam. Pristioidea
- Fam. Pristidae
- Fam. Torpedinidae
- Supérfam. Rajoidea
- Supérfam. Myliobatoidea
- Fam. Myliobatidae
- Fam. Trygonidae
- Fam. Hypolophidae
- Ordre Carcharhinida
- Fam. Palaeospinacidae
- Fam. Scyliorhinidae
- Fam. Triakidae
- Fam. Carcharhinidae
- Fam. Sphyrnidae
- Infraclasse Osteodonta
- Supérordre Ctenacanthi
- Ordre Ctenacanthida
- Ordre Tristychiida
- Supérordre Hybodonti
- Ordre Hybodontida
- Fam. Hybodontidae
- Fam. Ptychodontidae
- Supérordre Lamnae
- Ordre Orthacodontida
- Ordre Odontaspidida (=Lamniformes Berg excl. Orectolobidae)
- Supérfam. Odontaspidoidea
- Fam. Odontaspididae
- Fam. Jaekelotodontidae
- Fam. Otodontidae
- Fam. Carcharodontidae
- Fam. Cretoxyrhinidae
- Fam. Alopiidae
- Supérfam. Isuroidea
- Fam. Isuridae
- Fam. Lamiostomatidae
- Supérfam. Scapanorhynchidea
- Fam. Scapanorhynchidae
- Fam. Mitsukurinidae
- Supérfam. Anacoracoidea
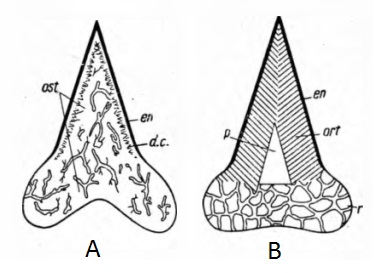
Fig. 1 : Coupes de dents d’Elasmobranches. A – d’un Ostéodonte. B – d’un Ortodonte. En – email, ost – osteodentine, ort – ortodentine, p – pulpe. D’après Glickman, 1964a.
L. S. Glickman a aussi développé une méthode d’étude de l’évolution des requins basée sur les dents, qui sont abondantes dans les roches sédimentaires depuis le Dévonien. Les descriptions de genres et d’espèces fondées sur un groupe de caractères arbitraires, très souvent utilisées en paléontologie, et jusqu’à il n’y a pas longtemps en biologie, se sont avérées inpraticables pour les requins, qui se ressemblent trop souvent entre eux. Beaucoup de requins de morphologie semblable ne sont pas unis par des liens de parenté, mais occupent la même niche écologique. Il est encore plus difficile d’étudier les requins éteints, dont seules les dents ont été préservées. Des tentatives de décrire les caractères des requins fossiles sont entreprises depuis longtemps, mais la grande quantité de caractères qu’on observe sur les dents de requins, ainsi que les liens de parenté incertains entre les requins éteints et actuels ont été les problèmes principaux des paléontologues. Glickman a démontré que les proportions de la couronne d’une dent de requin sont liées à sont type d’alimentation, et par conséquent à son mode de vie. Les dents de requins peuvent être approximativement divisées en quatre groupes: plurifonctionnelles (ударно-хватательные dans Glickman, 1964a), perforantes (колющие), coupantes ou déchireuses (режущие, рвущие), et broyeuses (дробящие). Les dents plurifonctionnelles ont la forme d’un cône, assez allongé et épais par rapport à sa largeur. Elles permettent de transpercer des proies, de briser des carapaces, ainsi que de déchirer, et caractérisent des requins adaptés à une grande variété de proies et d’habitats (exemple – le genre Lamna). Les dents perforantes sont longues et fines, et permettent uniquement d’attraper. Ce type de dent appartient aux requins vivant près des côtes et se nourrissant de poisson et de céphalopodes (exemple – le genre Odontaspis). Les dents coupantes sont larges et fines, les dents déchireuses ont par contre une couronne épaisse. Ce genre de dents permet de découper ou de déchirer des proies, et appartient aux superprédateurs (exemples - Isurus, Carcharodon, Squalicorax). Les dents broyeuses sont plates ou coniques, mais beaucoup plus courtes que les dents plurifonctionnelles, et sont caractéristiques des requins habitant près du fond marin, dont les proies sont des mollusques (autant bivalves que céphalopodes), les petits poissons et les crustacés (exemples - Squatina, Heterodontus). Les proportions des dents sont liées à de nombreux autres caractères (par exemples la forme et la disposition des nageoires), car elles ont une grande influence sur le mode de vie. Au cours de l’évolution, les requins s’adpatent à d’autres niches écologiques, ce qui entraîne des changements des proportions de la couronne en rapport avec la nouvelle fonction de la dent. Dans un deuxième temps, la fonctionnalité de la dent se perfectionne (p. ex. la dent s’allonge ou s’élargit encore plus), en même temps d’autres caractères secondaires apparaissent, liés à un manque de place dans la machoire, une surcharge de la dent etc. (caractères fréquents — épaississement de la base de la couronne pour renforcer la dent, atrophie des dents postérieures à cause de l’élargissement des dents antérieures et latérales). L’étude de l’évolution des requins est redue plus difficile par l’hétérodontie, c’est à dire que les dents anterieures et latérales, celles de la mâchoire supérieure et celle de l’infèrieure ont souvent des fonctions différentes, et par le fait que les dents de diverses positions peuvent évoluer à une vitesse différente. La méthode proposée par L. S. Glickman consiste à identifier les types de prédation des requins fossiles en se basant sur la morphologie fonctionnelle des dents, et à étudier les adaptations secondaires de l’appareil dentaire. En étudiant les dents d’âges différents à l’aide de la méthode de Glickman, il est possible de reconstruire l’évolution des genres et familles de requins, en se basant sur les régularités dans les changements de la géometrie des couronnes des dents, et sur les adaptations secondaires à ces changements[3, 5, 7 et autres].
L’étude de l’évolution des requins avec la méthode de Glickman à montré leur évolution rapide. L. S. Glickman a proposé d’utiliser les dents de requins en biostratigraphie — les dents de requins permettent d’identifier l’âge des roches avec précision, et les schémas biostratigraphiques basés sur les requins seraient utiles dans le monde entier, car la plupart des espèces de requins habitent toutes ou presque toutes les mers. La biostratigraphie à l’aide des dents de requins a plus tard été étudiée par R. A. Schwajeaite, V. I. Zhelezko et d’autres scientifiques.
- Glickman L. S., 1953 : Верхнемеловые позвоночные окрестностей Саратова. Предварительные данные. [Les vertébrés du Crétacé supérieur de la region de Saratov. Données préliminaires, en russe] // Учёные записки СГУ, 38:51-54.
- Glickman L. S., 1957b : О генетической связи Семейств Lamnidae и Odontaspidae и новых родах верхнемеловых ламнид [Du lien génétique des les familles Lamnidae et Odontaspidae et des nouveaux genres de lamnides du Crétacé supérieur, en russe] // Труды геол. музея им. А. П. Карпинского, 1:110-117.
- Glickman L. S., 1958b : О тэмпах эволюции ламноидных акул. [Du rythme de l’évolution des requins lamniformes, en russe] // Доклады АН СССР, 123(3):568-571.
- Glickman L. S., 1959 : Направление эволюционного развития и экология некоторых групп меловых эласмобранхий. [Le sens du développement évolutionnaire et l’écologie de certains groupes d’elasmobranches du Crétacé, en russe] // Труды 2й сессии Всесоюзного палеонтологического общества 226-234.
- Glickman L. S., 1964a :Акулы палеогена и их стратиграфическое значение. [Les requins du paléogène et leur valeur stratigraphique, en russe]. Наука.
- Glickman L. S., 1964b : Подкласс Elasmobranchii. Акуловые. [sous-classe Elasmobranchii. Requins, en russe]. Основы палеонтологии том 11: Бесчелюстные и рыбы 196-237, Наука.
- Glickman L. S., 1980 : Эволюция меловых и кайнозойских ламноидных акул. [Evolution des requins lamniformes du Crétacé et du Cénozoïque, en russe]. Наука.
- Popov, E. V., 2016 : An annotated bibliography of the soviet palaeoichtyologist Leonid Glickman (1929–2000) [en anglais] // Proceedings of the Zoological Institute RAS, 320(1):25–49.
- Popov E. V. et Glickman E. L., 2016 : The life and scientific heritage of Leonid Sergeyevich Glickman (1929-2000) [en russe]. Труды ЗИН РАН, 320(1):4-24.
{kind=link}
{kind=link}